Развитие нервной системы. Этапы развития нервной системы
Простейшие одноклеточные организмы не имеют нервной системы, регуляция жизнедеятельности у них происходит только за счёт гуморальных механизмов. При этом под действием какого-либо фактора внешней или внутренней среды увеличивается выработка регуляторных молекул, которые выделяются непосредственно во внутриклеточную жидкость (лат. «humor » – жидкость) и поступают к рабочей органелле путём диффузии. Только после этого формируется ответная реакция на причинный фактор. Разумеется, такой способ регуляции не является оперативным и точным, а, значит, ограничивает приспособительные возможности организма.
Нервная система, появившаяся у многоклеточных организмов, позволяет управлять системами организма более дифференцированно и с меньшими потерями времени на проведение командного сигнала (стимула). Поэтому у всех современных высокоорганизованных животных при единой нейрогуморальной регуляции функций организма ведущая роль принадлежит именно нервной системе. Филогенез нервной системы, то есть её эволюционное развитие (греч. «phylon » – род), предположительно, происходил в несколько этапов:
I этап – образование сетевидной нервной системы. На современном этапе эволюции такой тип нервной систем имеют кишечнополостные, например, гидра (рис. 12). Все нейроны у них являются мультиполярными и объединяются за счёт своих отростков в единую сеть, пронизывающую всё тело. При раздражении любой точки тела гидры возбуждается вся нервная система, вызывая движение всего тела. Эволюционным отголоском этого этапа у человека является сетевидное строение интрамуральной* нервной системы пищеварительного тракта (метасимпатической вегетативной нервной системы).
II этап – формирование узловой нервной системы связан с дальнейшей интеграцией организма и необходимостью централизованной переработки информации для ускорения этого процесса. На этом этапе произошла специализация нейронов и их сближение с образованием нервных узлов – центров. Отростки этих нейронов образовали нервы, идущие к рабочим органам. Централизация нервной системы привела к формированию рефлекторных дуг. Процесс централизации происходил двумя путями (рис. 13): с образованием радиальной (несимметричной) нервной системы (иглокожие, моллюски) и лестничной (симметричной) системы (например, плоские и круглые черви).
Радиальная нервная система, при которой все нервные ганглии сосредотачиваются в одном или двух-трёх местах, оказалась мало перспективной в эволюционном плане. Из животных, имеющих несимметричную ЦНС, только осьминоги достигли низшего уровня перцептивной психики, остальные же не поднялись выше сенсорной психики.
При формировании ЦНС лестничного типа (как, например, у планарий, см. рис. 13, А) ганглии формируются в каждом сегменте тела и соединяются между собой, а также с сегментами верхних и нижних уровней посредством продольных стволов. На переднем конце нервной системы развиваются нервные узлы, отвечающие за восприятие информации от передней части тела, которая в процессе движения первой и чаще сталкивается с новыми стимулами. В связи с этим головные ганглии беспозвоночных развиты сильнее остальных, являясь прообразом будущего головного мозга. Отражением этого этапа формирования ЦНС у человека является строение вегетативной нервной системы в виде параллельно идущих цепочек симпатических ганглиев.
III этапом является образование трубчатой нервной системы . Такая ЦНС впервые возникла у хордовых (ланцетник) в виде метамерной* нервной трубки с отходящими от неё сегментарными нервами ко всем сегментам туловища – туловищный мозг (рис. 14). Появление туловищного мозга связано с усложнением и совершенствованием движений, требующих координированного участия мышечных групп разных сегментов тела.
IV этап связан с образованием головного мозга . Этот процесс называется цефализацией (от греч. «encephalon » – головной мозг). Дальнейшая эволюция ЦНС связана с обособлением переднего отдела нервной трубки, что первоначально обусловлено развитием анализаторов, и приспособлением к разнообразным условиям обитания (рис. 15).
Филогенез головного мозга, согласно схеме Е.К. Сеппа и соавт. (1950), также проходит несколько этапов. На первом этапе цефализации из переднего отдела нервной трубки формируются три первичных пузыря . Развитие заднего пузыря (первичный задний , или ромбовидный мозг , rhombencephalon ) происходит у низших рыб в связи с совершенствованием слухового и вестибулярного анализаторов, воспринимающих звук и положение тела в пространстве (VIII пара головных нервов). Эти два вида анализаторов наиболее важны для ориентации в водной среде и являются, вероятно, эволюционно наиболее ранними. Так как на этом этапе эволюции наиболее развит задний мозг, в нём же закладываются и центры управления растительной жизнью, контролирующие важнейшие системы жизнеобеспечения организма – дыхательную, пищеварительную и систему кровообращения. Такая локализация сохраняется и у человека, у которого выше указанные центры располагаются в продолговатом мозге.
Задний мозг по мере развития делится на собственно задний мозг (metencephalon ), состоящий из моста и мозжечка, и продолговатый мозг (myelencephalon ), являющийся переходным между головным и спинным мозгом.
На втором этапе цефализации произошло развитие второго первичного пузыря (mesencephalon ) под влиянием формирующегося здесь зрительного анализатора; этот этап также начался ещё у рыб.
На третьем этапе цефализации формировался передний мозг (prosencephalon ), который впервые появился у амфибий и рептилий. Это было связано с выходом животных из водной среды в воздушную и усиленным развитием обонятельного анализатора, необходимого для обнаружения находящихся на расстоянии добычи и хищников. В последующем передний мозг разделился на промежуточный и конечный мозг (diencephalon et telencephalon ). Таламус стал интегрировать и координировать сенсорные функции организма, базальные ганглии конечного мозга стали отвечать за автоматизмы и инстинкты, а кора конечного мозга, сформировавшаяся изначально как часть обонятельного анализатора, со временем стала высшим интегративным центром, формирующим поведение на основе приобретённого опыта. Подробнее вопросы эволюции конечного мозга будут рассмотрены в Разделе 6.5.1.
V этап эволюции нервной системы – кортиколизация функций (от лат. «cortex » – кора) (рис. 16). Полушария большого мозга, возникшие у рыб в виде парных боковых выростов переднего мозга, первоначально выполняли только обонятельную функцию. Кора, сформировавшаяся на этом этапе и выполняющая функцию переработки обонятельной информации, называется древней корой (paleocortex , палеокортекс ). Она отличается малым числом слоёв нейронов (2–3) , что является признаком её примитивности. В процессе дальнейшего развития других отделов коры большого мозга древняя кора смещалась вниз и медиально. У разных видов она сохраняла свою функцию, но относительные её размеры уменьшались. У человека древняя кора представлена в области нижнемедиальной поверхности височной доли (переднее продырявленное вещество и смежные с ним участки), функционально она входит в лимбическую систему и отвечает за инстинктивные реакции (см. Раздел 6.5.2.1.2.).
Начиная с амфибий (см. рис. 16), происходит образование базальных ганглиев (структур полосатого тела) и так называемой старой коры (archicortex , архикортекс ) и повышается их значимость в формировании поведения. Базальные ганглии стали выполнять ту же функцию, что и архикортекс, значительно расширив диапазон и сложность автоматических, инстинктивных реакций.
Старая кора, как и древняя, состоит только из 2–3 слоёв нейронов. У амфибий и рептилий она занимает верхние участки больших полушарий. Однако, начиная с примитивных млекопитающих, по мере увеличения новой коры, она постепенно смещается на срединную поверхность полушарий. У человека этот вид коры находится в зубчатой извилине и гиппокампе.
Старая кора включена в лимбическую систему, в которую кроме неё входят таламус, миндалина, полосатое тело и древняя кора (см. Раздел 6.5.2.1.2.).
С образованием этой системы мозг приобретает новые функции – формирование эмоций и способность к примитивному научению на основе положительного или отрицательного подкрепления действий. Эмоции и ассоциативное научение значительно усложнили поведение млекопитающих и расширили их адаптационные возможности.
Дальнейшее совершенствование сложных форм поведения связано с формированием новой коры (neocortex , неокортекс). Нейроны новой коры впервые появляются у высших рептилий, однако, сильнее всего неокортекс развит у млекопитающих (см. рис. 16). У высших млекопитающих неокортекс покрывает увеличившиеся большие полушария, оттесняя вниз и медиально структуры древней и старой коры. Новая кора наиболее развита у человека, её площадь достигает 220 000 мм 2 , при этом две трети площади коры находится в её складках. Неокортекс становится центром обучения, памяти и интеллекта, может контролировать функции других отделов мозга, влияя на реализацию эмоциональных и инстинктивных форм поведения.
Таким образом, значимость кортиколизации функций заключается в том, что по мере своего развития кора конечного мозга берёт на себя роль высшего центра переработки информации и построения программ поведения. При этом корковые отделы анализаторов и корковые двигательные центры подчиняют себе нижележащие эволюционно более старые центры. В результате совершенствуется обработка информации, так как к интегративным возможностям подкорковых центров добавляются качественно новые возможности коры. Филогенетически старые сенсорные центры становятся переключающими центрами, осуществляющими начальную переработку информации, окончательная оценка которой будет произведена лишь в коре больших полушарий.
По такой же схеме строится и формирование поведения: инстинктивные, видоспецифические автоматические действия регулируются подкорковыми ядрами, а благоприобретенные компоненты поведения, вырабатывающиеся в течение всей жизни, формируются корой. Кора же может управлять центрами инстинктивных реакций, существенно расширяя при этом диапазон поведенческих реакций.
Кортиколизация функций увеличивается при переходе на более высокий уровень эволюционного развития и сопровождается увеличением площади коры и усилением её складчатости.
В эволюции нервная система претерпела несколько этапов развития, которые стали поворотными пунктами в качественной организации её деятельности. Эти этапы отличаются по количеству и видам нейрональных образований, синапсов, признакам их функциональной специализации, по образованию группировок нейронов, связанных между собой общностью функций. Выделяют три основных этапа структурной организации нервной системы: диффузный, узловой, трубчатый.
Диффузная нервная система наиболее древняя, имеется у кишечнополостных (гидра) животных. Такая нервная система характеризуется множественностью связей соседних элементов, что позволяет возбуждению свободно распространяться по нервной сети во все стороны.
Этот тип нервной системы обеспечивает широкую взаимозаменяемость и тем самым большую надёжность функционирования, однако эти реакции имеют неточный, расплывчатый характер.
Узловой тип нервной системы типичен для червей, моллюсков, ракообразных.
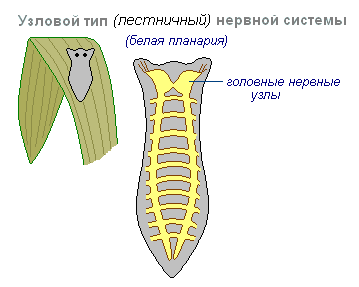
Он характерен тем, что связи нервных клеток организованы определённым образом, возбуждение проходит по жёстко определённым путям. Такая организация нервной системы оказывается более ранимой. Повреждение одного узла вызывает нарушение функций всего организма в целом, но она по своим качествам быстрее и точнее.
Трубчатая нервная система характерна для хордовых, она включает в себя черты диффузного и узлового типов. Нервная система высших животных взяла всё лучшее: высокую надёжность диффузного типа, точность, локальность быстроту организации реакций узлового типа.
Ведущая роль нервной системы
На первом этапе развития мира живых существ взаимодействие между простейшими организмами осуществлялось через водную среду первобытного океана, в которую поступали химические вещества, выделяемые ими. Первой древнейшей формой взаимодействия между клетками многоклеточных организм является химическое взаимодействие посредством продуктов обмена веществ, поступающих в жидкости организма. Такими продуктами обмена веществ, или метаболитами, являются продукты распада белков, углекислота и др. это — гуморальная передача влияний, гуморальный механизм корреляции, или связи между органами.
Гуморальная связь характеризуется следующими особенностями:
- отсутствием точного адреса, по которому направляется химическое вещество, поступающее в кровь или другие жидкости тела;
- химическое вещество распространяется медленно;
- химическое вещество действует в ничтожных количествах и обычно быстро разрушается или выводится из организма.
Гуморальные связи являются общими и для мира животных, и для мира растений. На определённой ступени развития мира животных в связи с появлением нервной системы образуется новая, нервная форма связей и регуляций, которая качественно отличает мир животных от мира растений. Чем выше по своему развитию организм животного, тем большую роль играет взаимодействие органов через нервную систему, которое обозначается как рефлекторное. У высших живых организмов нервная система регулирует гуморальные связи. В отличие от гуморальной связи нервная связь имеет точную направленность к определённому органу и даже группе клеток; связь осуществляется в сотни раз с большей скоростью, чем скорость распространения химических веществ. Переход от гуморальной связи к нервной сопровождался не уничтожением гуморальной связи между клетками тела, а подчинением нервным связям и возникновению нервно-гуморальным связям.
На следующем этапе развития живых существ появляются специальные органы — железы, в которых вырабатываются гормоны, образующиеся из поступающих в организм пищевых веществ. Основная функция нервной системы заключается как в регуляции деятельности отдельных органов между собой, так и во взаимодействии организма как единого целого с окружающей его внешней средой. Любое воздействие внешней среды на организм оказывается, прежде всего, на рецепторы (органы чувств) и осуществляется через посредство изменений, вызываемых внешней средой и нервной системой. По мере развития нервной системы высший её отдел — большие полушария головного мозга — становится «распорядителем и распределителем всей деятельности организма».
Строение нервной системы
Нервная система образована нервной тканью, которая состоит из огромного количества нейронов — нервная клетка с отростками.

Нервная система условно подразделяется на центральную и периферическую.

Центральная нервная система включает головной и спинной мозг, а периферическая нервная система - нервы, отходящие от них.

Головной и спинной мозг представляют собой совокупность нейронов. На поперечном разрезе мозга различают белое и серое вещество. Серое вещество состоит из нервных клеток, а белое - из нервных волокон, являющихся отростками нервных клеток. В различных отделах центральной нервной системы расположение белого и серого вещества неодинаково. В спинном мозге серое вещество находится внутри, а белое — снаружи, в головном же (большие полушария, мозжечок), наоборот — серое вещество — снаружи, белое — внутри. В различных отделах головного мозга имеются отдельные скопления нервных клеток (серого вещества), расположенные внутри белого вещества, - ядра . Скопления нервных клеток находятся и за пределами центральной нервной системы. Они называются узлами и относятся к периферической нервной системе.
Рефлекторная деятельность нервной системы
Основной формой деятельности нервной системы является рефлекс. Рефлекс - реакция организма на изменение внутренней или внешней среды, осуществляемая при участии центральной нервной системы в ответ на раздражение рецепторов.
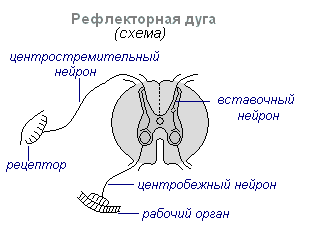
При всяком раздражении возбуждение с рецепторов передаётся по центростремительным нервным волокнам в центральную нервную систему, откуда через вставочный нейрон по центробежным волокнам оно идёт на периферию к тому или иному органу, деятельность которого изменяется. Весь этот путь через центральную нервную систему к рабочему органу, называется рефлекторной дугой образован обычно тремя нейронами: чувствительным, вставочным и двигательным. Рефлекс — сложный акт, в осуществлении которого принимает участие значительно большее количество нейронов. Возбуждение, попадая в центральную нервную систему, распространяется на многие отделы спинного мозга и доходит до головного. В результате взаимодействия многих нейронов осуществляется ответная реакция организма на раздражение.
Спинной мозг
Спинной мозг - тяж длиной около 45 см, диаметром 1 см, находится в канале позвоночника, покрыт тремя мозговыми оболочками: твёрдой, паутинной и мягкой (сосудистой).

Спинной мозг находится в позвоночном канале и представляет собой тяж, который вверху переходит в продолговатый мозг, а внизу заканчивается на уровне второго поясничного позвонка. Спинной мозг состоит из серого вещества, содержащего нервные клетки, и белого, состоящего из нервных волокон. Серое вещество расположено внутри спинного мозга и окружено со всех сторон белым веществом.

На поперечном разрезе серое вещество напоминает букву Н. В нём различают передние и задние рога, а также соединяющую перекладину, в центре которой находится узкий канал спинного мозга, содержащий спинномозговую жидкость. В грудном отделе выделяют боковые рога. В них заложены тела нейронов, иннервирующих внутренние органы. Белое вещество спинного мозга образовано нервными отростками. Короткие отростки соединяют участки спинного мозга, а длинные составляют проводниковый аппарат двусторонних связей с головным мозгом.

Спинной мозг имеет два утолщения - шейное и поясничное, от которых отходят нервы к верхним и нижним конечностям. От спинного мозга отходит 31 пара спинномозговых нервов. Каждый нерв начинается от спинного мозга двумя корешками — передним и задним. Задние корешки — чувствительные состоят из отростков центростремительных нейронов. Их тела расположены в спинномозговых узлах. Передние корешки — двигательные — являются отростками центробежных нейронов расположенных в сером веществе спинного мозга. В результате слияния переднего и заднего корешка образуется смешанный спинномозговой нерв. В спинном мозге сосредоточены центры, регулирующие наиболее простые рефлекторные акты. Основные функции спинного мозга - рефлекторная деятельность и проведение возбуждения.

В спинном мозге человека заложены рефлекторные центры мышц верхних и нижних конечностей, потоотделения и мочеиспускания. Функции проведения возбуждения заключается в том, что через спинной мозг проходят импульсы от головного мозга ко всем областям тела и обратно. По восходящим проводящим путям в головной мозг передаются центростемительные импульсы от органов (кожа, мышцы). По нисходящим путям центробежные импульсы передаются от головного мозга в спинной, затем на периферию, к органам. При повреждении проводящих путей наблюдается потеря чувствительности в различных участках тела, нарушение произвольных сокращений мышц и способности к движению.
Эволюция головного мозга позвоночных
Образование центральной нервной системы в виде нервной трубки впервые появляется у хордовых. У низших хордовых нервная трубка сохраняется в течение всей жизни, у высших - позвоночных - в стадии эмбриона на спинной стороне закладывается нервная пластинка, которая погружается под кожу и сворачивается в трубку. В эмбриональной стадии развития нервная трубка образует в передней части три вздутия — три мозговых пузыря, из которых развиваются отделы мозга: передний пузырь дает передний и промежуточный мозг, средний пузырь превращается в средний мозг, задний пузырь образует мозжечок и продолговатый мозг . Эти пять отделов мозга характерны для всех позвоночных животных.
Для низших позвоночных - рыб и земноводных - характерно преобладание среднего мозга над остальными отделами. У земноводных несколько увеличивается передний мозг и в крыше полушарий образуется тонкий слой нервных клеток - первичный мозговой свод, древняя кора. У рептилий значительно увеличивается передний мозг за счет скоплений нервных клеток. Большую часть крыши полушарий занимает древняя кора. Впервые у рептилий появляется зачаток новой коры. Полушария переднего мозга наползают на другие отделы, вследствие чего образуется изгиб в области промежуточного мозга. Начиная с древних рептилий, полушария головного мозга становятся самым большим отделом головного мозга.

В строении головного мозгаптиц и пресмыкающихся много общего. На крыше головного мозга - первичная кора, хорошо развит средний мозг. Однако у птиц по сравнению с рептилиями возрастают общая масса мозга и относительные размеры переднего мозга. Мозжечок крупный и имеет складчатое строение. У млекопитающих передний мозг достигает наибольшей величины и сложности. Большую часть мозгового вещества составляет новая кора, которая служит центром высшей нервной деятельности. Промежуточный и средний отделы мозга у млекопитающих невелики. Разрастающиеся полушария переднего мозга накрывают их и подминают под себя. У некоторых млекопитающих мозг гладкий, без борозд и извилин, но у большинства млекопитающих в коре мозга имеются борозды и извилины. Появление борозд и извилин происходит вследствие роста мозга при ограниченных размерах черепа. Дальнейший рост коры приводит к появлению складчатости в виде борозд и извилин.
Головной мозг
Если спинной мозг у всех позвоночных животных развит более или менее одинаково, то головной мозг существенно отличатся размерами и сложностью строения у разных животных. Особенно резкие изменения в ходе эволюции претерпевает передний мозг. У низших позвоночных передний мозг развит слабо. У рыб он представлен обонятельными долями и ядрами серого вещества в толще мозга. Интенсивное развитие переднего мозга связано с выходом животных на сушу. Он дифференцируется на промежуточный мозг и на два симметричных полушария, которые называются конечным мозгом . Серое вещество на поверхности переднего мозга (кора) впервые появляется у пресмыкающихся, развиваясь далее у птиц и особенно у млекопитающих. Действительно большими полушариями переднего мозга становятся только у птиц и млекопитающих. У последних они покрывают почти все другие отделы головного мозга.
Головной мозг расположен в полости черепа. В него входят ствол и конечный мозг (кора больших полушарий).

Ствол мозга состоит из продолговатого мозга, варолиева моста, среднего и промежуточного мозга.
Продолговатый мозг является непосредственным продолжением спинного мозга и расширяясь, переходит в задний мозг. Он в основном сохраняет форму и строение спинного мозга. В толще продолговатого мозга расположены скопления серого вещества — ядра черепно-мозговых нервов. В состав заднего моста входят мозжечок и варолиев мост . Мозжечок расположен над продолговатым мозгом и имеет сложное строение. На поверхности полушарий мозжечка серое вещество образует кору, а внутри мозжечка - его ядра. Как и спинной продолговатый мозг выполняет две функции: рефлекторную и проводниковую. Однако рефлексы продолговатого мозга более сложные. Это выражается в важном значении в регуляции сердечной деятельности, состоянии сосудов, дыхания, потоотделения. В продолговатом мозге расположены центры всех этих функций. Здесь же находятся центры жевания, сосания, глотания, отделения слюны и желудочного сока. Несмотря на малую величину (2,5–3 см), продолговатый мозг представляет собой жизненно важный отдел ЦНС. Повреждение его может стать причиной смерти вследствие прекращения дыхания и деятельности сердца. Проводниковая функция продолговатого мозга и варолиева моста заключается в передаче импульсов из спинного мозга в головной и обратно.
В среднем мозге расположены первичные (подкорковые) центры зрения и слуха, которые осуществляют рефлекторные ориентировочные реакции на световые и звуковые раздражения. Эти реакции выражаются в различных движениях туловища, головы и глаз в сторону раздражителей. Средний мозг состоит из ножек мозга и четверохолмия. Средний мозг регулирует и распределяет тонус (напряжение) скелетных мышц.
Промежуточный мозг состоит из двух отделов - таламус и гипоталамус , каждый из которых состоит из большого числа ядер зрительных бугров и подбугровой области. Через зрительные бугры центростремительные импульсы передаются к коре больших полушарий от всех рецепторов тела. Ни один центростремительный импульс, откуда бы он ни шёл, не может пройти к коре, минуя зрительные бугры. Таким образом, через промежуточный мозг осуществляется связь всех рецепторов с корой больших полушарий. В подбугровой области расположены центры, оказывающие влияние на обмен веществ, терморегуляцию и железы внутренней секреции.
Мозжечок находится позади продолговатого мозга. Он состоит из серого и белого вещества. Однако в отличие от спинного мозга и ствола серое вещество - кора - находится на поверхности мозжечка, а белое вещество расположено внутри, под корой. Мозжечок координирует движения, делает их чёткими и плавными, играет важную роль в сохранении равновесия тела в пространстве, а также оказывает влияние на тонус мышц. При поражении мозжечка у человека наблюдается падение тонуса мышц, расстройство движений и изменение походки, замедляется речь и т.д. Однако через некоторое время движения и мышечный тонус восстанавливаются благодаря тому, что неповреждённые участки центральной нервной системы берут на себя функции мозжечка.
Большие полушария - наиболее крупный и развитый отдел головного мозга. У человека они образуют основную массу головного мозга и по всей своей поверхности покрыты корой. Серое вещество покрывает полушария снаружи и образует кору головного мозга. Кора полушарий человека имеет толщину от 2 до 4 мм и слагается из 6–8 слоёв, образованных 14–16 млрд. клеток, различных по форме, величине и выполняемым функциям. Под корой находится белое вещество. Оно состоит из нервных волокон, связывающих кору с расположенными ниже отделами центральной нервной системы и отдельные доли полушарий между собой.
Кора головного мозга имеет извилины, разделённые бороздами, которые значительно увеличивают её поверхность. Три самые глубокие борозды делят полушария на доли. В каждом полушарии различают четыре доли: лобную, теменную, височную, затылочную . Возбуждение разных рецепторов поступают в соответствующие воспринимающие участки коры, называемые зонами , и отсюда передаются к определённому органу, побуждая его к действию. В коре выделяют следующие зоны. Слуховая зона расположена в височной доле, воспринимает импульсы от слуховых рецепторов.

Зрительная зона лежит в затылочной области. Сюда поступают импульсы от рецепторов глаза.

Обонятельная зона находится на внутренней поверхности височной доли и связана с рецепторами носовой полости.

Чувствительно-двигательная зона расположена в лобной и теменной долях. В этой зоне находятся главные центры движения ног, туловища, рук, шеи, языка и губ. Здесь же лежит и центр речи.
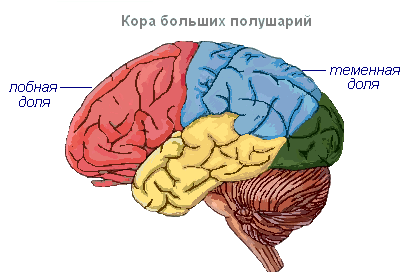
Полушария головного мозга - это высший отдел центральной нервной системы, контролирующий работу всех органов у млекопитающих. Значение больших полушарий у человека заключается ещё и в том, что они представляют собой материальную основу психической деятельности. И.П.Павлов показал, что в основе психической деятельности лежат физиологические процессы, происходящие в коре головного мозга. Мышление связано с деятельностью всей коры головного мозга, а не только с функцией отдельных её областей.
| Отдел головного мозга | Функции | |
| Продолговатый мозг | Проводниковая | Связь спинного и вышележащих отделов головного мозга. |
| Рефлекторная | Регуляция деятельности дыхательной, сердечно-сосудистой, пищеварительной систем:
|
|
| Варолиев мост | Проводниковая | Соединяет полушария мозжечка между собой и с корой больших полушарий головного мозга. |
| Мозжечок | Координационная | Координация произвольных движений и сохранение положения тела в пространстве. Регуляция мышечного тонуса и равновесия |
| Средний мозг | Проводниковая | Ориентировочные рефлексы на зрительные, звуковые раздражители (повороты головы и туловища ). |
| Рефлекторная |
|
|
| Промежуточный мозг | таламус
гипоталамус
|
|
Кора больших полушарий
Поверхность коры больших полушарий у человека составляет около 1500 см 2 , что во много раз превышает внутреннюю поверхность черепа. Такая большая поверхность коры образовалась благодаря развитию большого количества борозд и извилин, в результате чего большая часть коры (около 70%) сосредоточена в бороздах. Самые большие борозды больших полушарий - центральная , которая проходит поперёк обоих полушарий, и височная , отделяющая височную долю от остальных. Кора больших полушарий, несмотря на малую толщину (1,5–3 мм) имеет очень сложное строение. В ней насчитывают шесть основных слоёв, которые отличаются строением, формой и размерами нейронов и связями. В коре находятся центры всех чувствительных (рецепторных) систем, представительства всех органов и частей тела. В связи с этим к коре подходят центростремительные нервные импульсы от всех внутренних органов или частей тела, и она может управлять их работой. Через кору больших полушарий происходит замыкание условных рефлексов, посредством которых организм постоянно, в течение всей жизни очень точно приспосабливается к изменчивым условиям существования, к окружающей среде.
Среди беспозвоночных наиболее примитивный тип нервной системы в виде диффузной нервной сети встречается у кишечнополостных (см. рис. 1.2). Их нервная сеть представляет собой скопление мультиполярных и биполярных нейрон
ов
, отростки которых могут перекрещиваться, прилегать друг к другу и лишены функциональной дифференциации на аксон
ы
и дендрит
ы
. Диффузная нервная сеть не разделена на центральный и периферический отделы и может быть локализована в эктодерме и энтодерме.
Эпидермальные нервные сплетения, напоминающие нервные сети кишечнополостных, могут быть обнаружены и у более высоко организованных беспозвоночных (плоские и кольчатые черви), однако здесь они занимают подчиненное положение по отношению к ЦНС, которая выделяется как самостоятельный отдел.
...Ганглионизация нервных элементов получает дальнейшее развитие у высших беспозвоночных, кольчатых червей, моллюсков и членистоногих. У большинства кольчатых червей брюшные стволы ганглионизированы таким образом, что в каждом сегменте тела формируется по одной паре ганглиев, соединенных коннективами с другой парой, расположенной в соседнем сегменте.
...Эволюция нервной системы беспозвоночных идет не только по пути концентрации нервных элементов, но и в направлении усложнения структурных взаимоотношений в пределах ганглиев. Не случайно брюшную нервную цепочку сравнивают со спинным мозгом позвоночных животных. Как и в спинном мозгу, в ганглиях обнаруживается поверхностное расположение проводящих путей, дифференциация нейропиля на моторную, чувствительную и ассоциативные области.
...Прогрессивное развитие мозга у головоногих моллюсков и насекомых создает предпосылку для возникновения своеобразной иерархии командных систем управления поведением. Низший уровень интеграции в сегментарных ганглиях насекомых и в подглоточной массе мозга моллюсков служит основой для автономной деятельности и координации элементарных двигательных актов. В то же время мозг представляет собой следующий, более высокий уровень интеграции, где могут осуществляться межанализаторный синтез и оценка биологической значим
ости
информации. На основе этих процессов формируются нисходящие команды, обеспечивающие вариантность запуска нейрон
ов
сегментарных центров. Очевидно, взаимодействие двух уровней интеграции лежит в основе пластичности поведения высших беспозвоночных, включающего врожденные и приобретенные реакции.
...Нервная система позвоночных закладывается в виде сплошной нервной трубки, которая в процессе онто— и филогенеза дифференцируется на различные отделы и является также источником периферических симпатических, парасимпатических и метасимпатических нервных узлов. У наиболее древних хордовых (бесчерепных) головной мозг отсутствует, и нервная трубка представлена в малодифференцированном состоянии.
...В ходе дальнейшей эволюции наблюдается перемещение некоторых функций и систем интеграции из спинного мозга в головной — процесс энцефализации, который был рассмотрен выше на примере беспозвоночных животных. В период филогенетического развития от уровня бесчерепных до уровня круглоротых формируется головной мозг как надстройка над системами дистантной рецепции.
...Передний мозг круглоротых длительное время считали чисто обонятельным. Однако исследования недавнего времени показали, что обонятельные входы в передний мозг не являются единственными, а дополняются сенсорными входами других модальностей. Очевидно, уже на ранних этапах филогенеза позвоночных передний мозг начинает участвовать в переработке информации и управлении поведением. Вместе с тем энцефализация как маги
ст
ральное направление развития мозга не исключает эволюционных преобразований в спинном мозгу круглоротых. В отличие от бесчерепных нейрон
ы
кожной чувствительности выделяются из спинного мозга и концентрируются в спинномозговой ганглий. Наблюдается совершенствование проводниковой части спинного мозга. Проводящие волокна боковых столбов имеют контакты с мощной дендрит
ной
сетью мотонейрон
ов. Формируются нисходящие связи головного мозга со спинным через мюллеровские волокна — гигантские аксон
ы
клеток, лежащих в среднем и продолговатом мозгу.
...Наиболее существенные в эволюционном плане изменения происходят в промежуточном мозгу амфибий. Здесь обособляется таламус (зрительный бугор), дифференцируются структурированные ядра (наружное коленчатое тело) и восходящие пути, связывающие зрительный бугор с корой (таламокортикальный путь).
В полушариях переднего мозга происходит дальнейшая дифференциация зачатков старой и древней коры. В старой коре (археокортексе) обнаруживаются звездчатые и пирамидные клетки. В промежутке между старой и древней корой появляется полоска плаща, которая является предтечей новой коры (неокортекса).
В целом развитие переднего мозга создает предпосылки для перехода от свойственной рыбам мезэнцефалоцеребреллярной системы интеграции к диэнцефалотелэнцефалъной, где ведущим отделом становится передний мозг, а таламус промежуточного мозга превращается в коллектор всех афферентных сигналов. В полной мере эта система интеграции представлена в зауропсидном типе мозга у рептилий и знаменует собой следующий этап морфофункциональной эволюции мозга.
Развитие таламокортикальной системы связей у рептилий приводит к формированию новых проводящих путей, как бы подтягивающихся к филогенетически молодым формациям мозга.
В боковых столбах спинного мозга рептилий обособляется восходящий спинно—таламический путь, который проводит к головному мозгу информацию о температурной и болевой чувствительности. Здесь же в боковых столбах формируется новый нисходящий путь — красно—ядерно—спинномозговой (Монакова). Он связывает мотонейрон
ы спинного мозга с красным ядром среднего мозга, которое включено в древнюю экстрапирамидную систему двигательной регуляции. Эта многозвенная система объединяет влияние переднего мозга, мозжечка, ретикулярной формации ствола, ядер вестибулярного комплекса и координирует двигательную активность. У рептилий, как истинно наземных животных, возрастает роль зрительной и акустической информации, возникает необходимость сопоставления этой информации с обонятельной и вкусовой. В соответствии с этими биологическими изменениями в стволовой части мозга рептилий происходит целый ряд структурных изменений. В продолговатом мозгу дифференцируются слуховые ядра, помимо улиткового ядра появляется угловое, связанное со средним мозгом. В среднем мозгу двухолмие преобразуется в четверохолмие, в ростральных холмах которого находятся акустические центры.
Наблюдается дальнейшая дифференциация связей крыши среднего мозга с таламусом, который является как бы преддверия входа в кору всех восходящих сенсорных путей. В самом таламусе происходит дальнейшее обособление ядерных структур и установление между ними специализированных связей.
...У млекопитающих развитие переднего мозга сопровождалось бурным ростом новой коры, находящейся в тесной функциональной связи с таламусом промежуточного мозга. В коре закладываются эфферентные пирамидные клетки, посылающие свои длинные аксон
ы
к мотонейрон
ам спинного мозга.
Таким образом, наряду с многозвенной экстрапирамидной системой появляются прямые пирамидные пути, которые обеспечивают непосредственный контроль над двигательными актами. Корковая регуляция движений у млекопитающих приводит к развитию филогенетически наиболее молодой части мозжечка — передней части задних долей полушарий, или неоцеребеллума. Неоцеребеллум приобретает двусторонние связи с новой корой.
Рост новой коры у млекопитающих происходит настолько интенсивно, что старая и древняя кора оттесняются в медиальном направлении к мозговой перегородке. Бурный рост коры компенсируется формированием складчатости. У наиболее низко организованных однопроходных (утконос) на поверхности полушария закладываются первые две постоянные борозды, остальная же поверхность остается гладкой (лиссэнцефалический тип коры).
Как показали нейрофизиологические исследования, мозг однопроходных и сумчатых млекопитающих лишен еще соединяющего полушария мозолистого тела и характеризуется перекрытием сенсорных проекций в новой коре. Четкая локализация моторных, зрительных и слуховых проекций здесь отсутствует.
У плацентарных, млекопитающих (насекомоядных и грызунов) отмечается развитие более четкой локализации проекционных зон в коре. Наряду с проекционными зонами в новой коре формируются ассоциативные зоны, однако границы первых и вторых могут перекрываться. Мозг насекомоядных и грызунов характеризуется наличием мозолистого тела и дальнейшим увеличением общей площади новой коры, развитием борозд и извилин (гирэнцефалический тип коры).
В процессе параллельно—адаптивн
ой эволюции у хищных млекопитающих появляются теменные и лобные ассоциативные поля, ответственные за оценку биологически значим
ой
информации, мотивацию поведения и программирование сложных поведенческих актов. Наблюдается дальнейшее развитие складчатости новой коры.
И наконец, приматы демонстрируют наиболее высокий уровень организации коры головного мозга. Кора приматов характеризуется шестислойностью, отсутствием перекрытия ассоциативных и проекционных зон. У приматов формируются связи между фронтальными и теменными ассоциативными полями и, таким образом, возникает целостная интегративная система больших полушарий.
- 1)Дорзальная индукция или Первичная нейруляция - период 3-4 недели гестации;
- 2)Вентральная индукция - период 5-6 недели гестации;
- 3)Нейрональная пролиферация - период 2-4 месяца гестации;
- 4)Миграция - период 3-5 месяца гестации;
- 5)Организация - период 6-9 месяца развития плода;
- 6)Миелинизация - занимает период от момента рождения и в последующем периоде постнатальной адаптации.
В первом триместре беременности протекают такие этапы развития нервной системы плода:
Дорзальная индукция или Первичная нейруляция - в связи с индивидуальными особенностями развития может варьировать по времени, но всегда придерживается 3-4 неделе (18-27 день после зачатия) гестации. В этот период происходит образование нервной пластинки, которая после смыкания ее краев превращается в нервную трубку (4-7 неделя гестации).
Вентральная индукция - этот этап формирования нервной системы плода достигает своего пика на 5-6 неделе гестации. В этот период у нервной трубки появляются 3 расширенных полости (на переднем ее конце), из которых после формируются:
из 1-й (краниальной полости) - головной мозг;
из 2-й и 3-й полости - спинной мозг.
Вследствие деления на три пузыря, нервная система развивается дальше и зачаток головного мозга плода из трех пузырей превращается в пять путем деления.
Из переднего мозга образуется - конечный мозг и межуточный мозг.
Из заднего мозгового пузыря - закладка мозжечка и продолговатого мозга.
В первый триместр беременности также проходит частично нейрональная пролиферация.
Спинной мозг развивается быстрее, чем головной, и, следовательно, функционировать начинает также быстрее, отчего играет более важную роль на начальных этапах развития плода.
Но в первом триместре беременности особое внимание заслуживает процесс развития вестибулярного анализатора. Он является высокоспециализированным анализатором, который отвечает у плода за восприятие перемещения в пространстве и ощущение изменения положения. Этот анализатор формируется уже на 7 неделе внутриутробного развития (раньше других анализаторов!), а к 12-той неделе к нему уже подходят нервные волокна. Миелинизация нервных волокон начинается к моменту появления у плода первых движений - на 14 - неделе гестации. Но для проведения импульсов от вестибулярных ядер к двигательным клеткам передних рогов спинного мозга необходимо быть миелинизированным вестибуло - спинальному тракту. Его миелинизация происходит через 1-2 недели (15 - 16 неделя гестации).
Поэтому, благодаря раннему формированию вестибулярного рефлекса, при перемещении беременной женщины в пространстве плод перемещается в полости матки. Вместе с этим, перемещение плода в пространстве является «раздражающим» фактором для вестибулярного рецептора, который посылает импульсы для дальнейшего развития нервной системы плода.
Нарушения развития плода от воздействия различных факторов в этот период ведет к нарушениям вестибулярного аппарата у новорожденного ребенка.
До 2-го месяца гестации плод имеет гладкую поверхность головного мозга, покрытую эпендимным слоем, состоящим из медуллобластов. Ко 2 - му месяцу внутриутробного развития начинает формироваться кора головного мозга путем миграции нейробластов в вышележащий краевой слой, и, таким образом, формируя закладку серого вещества головного мозга.
Все неблагоприятные факторы воздействия в первый триместр развития нервной системы плода приводят к тяжелым и, в большинстве случаев, необратимым нарушениям функционирования и дальнейшего формирования нервной системы плода.
Второй триместр беременности.
Если в первом триместре беременности происходит основная закладка нервной системы, то во втором триместре происходит ее интенсивное развитие.
Нейрональная пролиферация является основным процессом онтогенеза.
На этом этапе развития возникает физиологическая водянка пузырей головного мозга. Это происходит из-за того, что спинномозговая жидкость, поступая в мозговые пузыри, расширяет их.
К концу 5-го месяца гестации образуются все основные борозды головного мозга, а также появляются отверстия Люшка, через которые спинномозговая жидкость выходит на наружную поверхность мозга и омывает его.
В течение 4 - 5 месяца развития мозга интенсивно развивается мозжечок. Он приобретает характерную ему извилистость, и делиться поперек, образуя свои основные части: переднюю, заднюю и фолликуло-нодулярные доли.
Также во втором триместре беременности проходит этап миграции клеток (5 месяц), в результате которого появляется зональность. Головной мозг плода становится более похож на головной мозг взрослого ребенка.
При воздействии неблагоприятных факторов на плод во второй период беременности, возникают нарушения, которые совместимы с жизнью, так как закладка нервной систему прошла в первом триместре. На этом этапе нарушения связанны с недоразвитием структур мозга.
Третий триместр беременности.
В этот период происходит организация и миелинизация структур головного мозга. Борозды и извилины в своем развитии подходят к завершающему этапу (7 - 8 месяц гестации).
Под этапом организации нервных структур понимают морфологическую дифференцировку и возникновение специфических нейронов. В связи с развитием цитоплазмы клеток и увеличения внутриклеточных органелл, происходит увеличение образования продуктов обмена, которые необходимы для развития нервных структур: белки, ферменты, гликолипиды, медиаторы и др. Параллельно с этими процессами протекает образование аксонов и дендритов для обеспечения синоптических контактов между нейронами.
Миелинизация нервных структур начинается с 4-5 месяца гестации и заканчивается к концу первого, началу второго года жизни ребенка, когда ребенок начинает ходить.
При воздействии неблагоприятных факторов в третьем триместре беременности, а также в течение первого года жизни, когда заканчиваются процессы миелинизации пирамидных путей, серьезных нарушений не возникает. Возможны легкие изменения структуры, которые определяются только при гистологическом исследовании.
Развитие ликвора и кровеносной системы головного и спинного мозга.
В первом триместре беременности (1 - 2 месяц гестации), когда происходит образование пяти мозговых пузырей, происходит образование сосудистых сплетений в полости первого, второго и пятого мозгового пузыря. Эти сплетения начинают секретировать высококонцентрированный ликвор, который является, по сути, питательной средой из-за большого содержания в своем составе белка и гликогена (превышает в 20 раз в отличие от взрослых). Ликвор - в этом периоде является основным источником питательных веществ для развития структур нервной системы.
Пока развитие мозговых структур поддерживает ликвор, на 3 - 4 неделе гестации образуются первые сосуды кровеносной системы, которые расположены в мягко-паутинной оболочке. Изначально содержание кислорода в артериях очень низкое, но в течение с 1 - го по 2 - й месяц внутриутробного развития кровеносная система приобретает более зрелый вид. И на втором месяце гестации кровеносные сосуды начинают врастать в мозговое вещество, образуя кровеносную сеть.
К 5 - му месяцу развития нервной системы появляются передняя, средняя и задняя мозговые артерии, которые соединены между собой анастомозами, и представляют собой завершенную структуру мозга.
Кровоснабжение спинного мозга происходит из большего количества источников, чем у головного мозга. Кровь к спинному мозгу поступает из двух позвоночных артерий, которые разветвляются на три артериальных тракта, которые, в свою очередь, идут вдоль всего спинного мозга, питая его. Передние рога получают большее количество питательных веществ.
Венозная система исключает образование коллатералей и является более изолированной, что способствует быстрому выведению конечных продуктов обмена по центральным венам на поверхность спинного мозга и выведением в венозные сплетения позвоночника.
Особенностью кровоснабжения третьего, четвертого и боковых желудочков у плода является более широкий размер капилляров, которые проходят в этих структурах. Это ведет к замедленному току крови, что способствует более интенсивному питанию.
| Возраст зародыша (недели) | Развитие нервной системы |
| 2,5 | Намечается нервная бороздка |
| 3.5 | Образуется нервная трубка и нервные тяжи |
| Образуются 3 мозговых пузыря; формируются нервы и ганглии | |
| Формируются 5 мозговых пузырей | |
| Намечаются мозговые оболочки | |
| Полушария мозга достигают большого размера | |
| В коре появляются типичные нейроны | |
| Формируется внутренняя структура спинного мозга | |
| Формируются общие структурные черты головного мозга; начинается дифференцировка клеток нейроглии | |
| Различимы доли головного мозга | |
| 20-40 | Начинается миелинизация спинного мозга (20 неделя), появляются слои коры (25 недель), формируются борозды и извилины (28-30 недель), начинается миелинизация головного мозга (36-40 недель) |
Таким образом, развитие головного мозга в пренатальный период происходит непрерывно и параллельно, однако характеризуется гетерохронией: скорость роста и развития филогенетически более древних образований больше, чем филогенетически более молодых образований.
Ведущую роль в росте и развитии нервной системы во внутриутробный период играют генетические факторы. Вес мозга новорожденного в среднем составляет примерно 350 г.
Морфо-функциональное созревание нервной системы продолжается в постнатальный период. Уже к концу первого года жизни вес мозга достигает 1000 г, тогда как у взрослого человека вес мозга составляет в среднем — 1400 г. Следовательно, основное прибавление массы мозга приходится на первый год жизни ребенка.
Увеличение массы мозга в постнатальный период происходит в основном за счет увеличения количества глиальных клеток. Количество нейронов не увеличивается, так как они теряют способность делиться уже в пренатальном периоде. Общая плотность нейронов (количество клеток в единице объема) уменьшается за счет роста сомы и отростков. У дендритов увеличивается количество ветвлений.
В постнатальном периоде продолжается также миелинизация нервных волокон как в центральной нервной системе, так и нервных волокон, входящих в состав периферических нервов (черепно-мозговых и спинномозговых.).
Рост спинномозговых нервов связан с развитием опорно-двигательного аппарата и формированием нервно-мышечных синапсов, а рост черепно-мозговых нервов с созреванием органов чувств.
Таким образом, если в пренатальном периоде развитие нервной системы происходит под контролем генотипа и практически не зависит от влияния внешней окружающей среды, то в постанатальном периоде все большую роль приобретают внешние стимулы. Раздражение рецепторов вызывает афферентные потоки импульсов, которые стимулируют морфо-функциональное созревание мозга.
Под влиянием афферентных импульсов на дендритах корковых нейронов образуются шипики — выросты, представляющие собой особые постсинаптические мембраны. Чем больше шипиков, тем больше синапсов и тем большее участие принимает нейрон в обработке информации.
На протяжении всего постнатального онтогенеза вплоть до пубертатного периоде также как и в пренатальный период развитие мозга происходит гетерохронно. Так, окончательное созревание спинного мозга происходит раньше, чем головного мозга. Развитие стволовых и подкорковых структур, раньше, чем корковых, рост и развитие возбудительных нейронов обгоняет рост и развитие тормозных нейронов. Это общие биологические закономерности роста и развития нервной системы.
Морфологическое созревание нервной системы коррелирует с особенностями ее функционирования на каждом этапе онтогенеза. Так, более раннее дифференцирование возбудительных нейронов по сравнению с тормозными нейронами обеспечивает преобладание мышечного тонуса сгибателей над тонусом разгибателей. Руки и ноги плода находятся в согнутом положении — это обуславливает позу, обеспечивающую минимальный объем, благодаря чему плод занимает меньшее место в матке.
Совершенствование координации движений, связанных с формированием нервных волокон, происходит на протяжении всего дошкольного и школьного периодов, что проявляется в последовательном освоении позы сидения, стояния, ходьбы, письма и т.д.
Увеличение скорости движений обуславливается в основном процессами миелинизации периферических нервных волокон и увеличения скорости проведения возбуждения нервных импульсов.
Более раннее созревание подкорковых структур по сравнению с корковыми, многие из которых входят в состав лимбической структуры, обуславливают особенности эмоционального развития детей (большая интенсивность эмоций, неумение их сдерживать связана с незрелостью коры и ее слабым тормозным влиянием).
В пожилом и старческом возрасте происходят анатомические и гистологические изменения мозга. Часто происходит атрофия коры лобной и верхней теменной долей. Борозды становятся шире, желудочки мозга увеличиваются, объем белого вещества уменьшается. Происходит утолщение мозговых оболочек.
С возрастом нейроны уменьшаются в размерах, при этом количество ядер в клетках может увеличиться. В нейронах уменьшается также содержание РНК, необходимой для синтеза белков и ферментов. Это ухудшает трофические функции нейронов. Высказывается предположение, что такие нейроны быстрее утомляются.
В старческом возрасте нарушается также кровоснабжение мозга, стенки кровеносных сосудов утолщаются и на них откладываются холестериновые бляшки (атеросклероз). Это также ухудшает деятельность нервной системы.
ЛИТЕРАТУРА
Атлас “Нервная система человека”. Сост. В.М. Асташев. М., 1997.
Блюм Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум и поведение. М.: Мир, 1988.
Борзяк Э.И., Бочаров В.Я., Сапина М.Р. Анатомия человека. — М.: Медицина, 1993. Т.2. 2-е изд., перераб. и доп.
Загорская В.Н., Попова Н.П. Анатомия нервной системы. Программа курса. МОСУ, М., 1995.
Кишш-Сентаготаи. Анатомический атлас человеческого тела. — Будапешт, 1972. 45-е изд. Т. 3.
Курепина М.М., Воккен Г.Г. Анатомия человека. — М.: Просвещение, 1997. Атлас. Изд.2-е.
Крылова Н.В., Искренко И.А. Мозг и проводящие пути (Анатомия человека в схемах и рисунках). М.: Изд-во Российского университета дружбы народов, 1998.
Мозг. Пер. с англ. Под ред. Симонова П.В. — М.: Мир, 1982.
Морфология человека. Под ред. Б.А. Никитюка, В.П. Чтецова. — М.: Изд-во МГУ, 1990. С. 252-290.
Привес М.Г., Лысенков Н.К., Бушкович В.И. Анатомия человека. — Л.: Медицина, 1968. С. 573-731.
Савельев С.В. Стереоскопический атлас мозга человека. М., 1996.
Сапин М.Р., Билич Г.Л. Анатомия человека. — М.: Высшая школа, 1989.
Синельников Р.Д. Атлас анатомии человека. — М.: Медицина, 1996. 6-е изд. Т. 4.
Шаде Дж., Форд Д. Основы неврологии. — М.: Мир, 1982.
РАЗДЕЛ I. ЦИТОЛОГИЧЕСКИЕ И ГИСТОЛОГИЧЕСКИЕ ХАРАКТЕРИСТИКИ НЕРВНОЙ СИСТЕМЫ 3
РАЗДЕЛ II. СТРОЕНИЕ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ. 30
РАЗДЕЛ III. ГОЛОВНОЙ МОЗГ………………………………………………………… 46
РАЗДЕЛ IV. РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ……………………………. 92
ЛИТЕРАТУРА…………………………………………………………………………………….. 102
Лицензия на издательскую деятельность:
серия ИД № 00865, дата регистрации: 25.01.2000 года
Адрес издательства: 109383, г. Москва, ул. Шоссейная, 86
Социально-технологический институт МГУС
Ткань — это совокупность клеток и межклеточного вещества, сходных по строению, происхождению и выполняемым функциям.
2 Некоторые анатомы продолговатый мозг не включают в задний мозг, а выделяют его в качестве самостоятельного отдела.
РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ В ОНТОГЕНЕЗЕ
Онтогенез, или индивидуальное развитие организма, делится на два периода: пренатальный (внутриутробный) и постнатальный (после рождения).
Первый продолжается от момента зачатия и формирования зиготы до рождения; второй – от момента рождения и до смерти.
Пренатальный период в свою очередь подразделяется на три периода: начальный, зародышевый и плодный.
Начальный (предимплантационный) период у человека охватывает первую неделю развития (с момента оплодотворения до имплантации в слизистую оболочку матки). Зародышевый (предплодный, эмбриональный) период – от начала второй недели до конца восьмой недели (с момента имплантации до завершения закладки органов).
Плодный (фетальный) период начинается с девятой недели и длится до рождения. В это время происходит усиленный рост организма.
Постнатальный период онтогенеза подразделяют на одиннадцать периодов: 1-й – 10-й день – новорожденные; 10-й день – 1 год – грудной возраст; 1–3 года – раннее детство; 4–7 лет – первое детство; 8–12 лет – второе детство; 13–16 лет – подростковый период; 17–21 год – юношеский возраст; 22–35 лет – первый зрелый возраст; 36–60 лет – второй зрелый возраст; 61–74 года– пожилой возраст; с 75 лет – старческий возраст, после 90 лет – долгожители.
Завершается онтогенез естественной смертью.
Пренатальный период онтогенеза начинается с момента слияния мужских и женских половых клеток и образования зиготы. Зигота последовательно делится, образуя шаровидную бластулу. На стадии бластулы идет дальнейшее дробление и образование первичной полости – бластоцеля.
Затем начинается процесс гаструляции, в результате которого происходит перемещение клеток различными способами в бластоцель, с образованием двухслойного зародыша.
Наружный слой клеток называется эктодерма , внутренний – энтодерма. Внутри образуется полость первичной кишки – гастроцель.
Это стадия гаструлы. На стадии нейрулы образуются нервная трубка , хорда , сомиты и другие эмбриональные зачатки.
Зачаток нервной системы начинает развиваться еще в конце стадии гаструлы.
Рис. 16. Закладка нервной трубки (схематичное изображение и вид на поперечном срезе):
А–А’– уровень поперечного среза; а – начальный этап погружения медуллярной пластинки и формирования нервной трубки: 1 – нервная трубка; 2 – ганглиозная пластина; 3 – сомит; б – завершение образования нервной трубки и погружение ее внутрь зародыша: 4 – эктодерма; 5 – центральный канал; 6 – белое вещество спинного мозга; 7 – серое вещество спинного мозга; 8 – закладка спинного мозга; 9 – закладка головного мозга
Клеточный материал эктодермы, расположенный на дорсальной поверхности зародыша, утолщается, образуя медуллярную пластинку (рис.
17, 2 ). Эта пластинка ограничивается с боков медуллярными валиками. Дробление клеток медуллярной пластинки (медуллобластов) и медуллярных валиков приводит к изгибанию пластинки в желоб, а затем к смыканию краев желоба и образованию медуллярной трубки (рис. 16а, 1 ). При соединении медуллярных валиков образуется ганглиозная пластина, которая затем делится на ганглиозные валики.

17. Пренатальное развитие нервной системы человека:
1 – нервный гребень; 2 – нервная пластина; 3 – нервная трубка; 4 – эктодерма; 5 – средний мозг; 6 – спинной мозг; 7 – спинномозговые нервы; 8 – глазной пузырек; 9 – передний мозг; 10 – промежуточный мозг; 11 – мост; 12 – мозжечок; 13 – конечный мозг
Одновременно происходит погружение нервной трубки внутрь зародыша (рис.
16в; 17, 3 ).
Однородные первичные клетки стенки медуллярной трубки – медуллобласты – дифференцируются на первичные нервные клетки (нейробласты) и исходные клетки нейроглии (спонгиобласты).
Клетки внутреннего, прилежащего к полости трубки, слоя медуллобластов превращаются в эпендимные, которые выстилают просвет полостей мозга. Все первичные клетки активно делятся, увеличивая толщину стенки мозговой трубки и уменьшая просвет нервного канала. Нейробласты дифференцируются на нейроны, спонгиобласты – на астроциты и олигодендроциты, эпендимные – на эпендимоциты (на этом этапе онтогенеза клетки эпендимы могут образовывать нейробласты и спонгиобласты).
При дифференцировке нейробластов отростки удлиняются и превращаются в дендриты и аксон, которые на данном этапе лишены миелиновых оболочек.
Миелинизация начинается с пятого месяца пренатального развития и полностью завершается лишь в возрасте 5–7 лет. На пятом же месяце появляются синапсы. Миелиновая оболочка формируется в пределах ЦНС олигодендроцитами, а в периферической нервной системе – Шванновскими клетками.
В процессе эмбрионального развития формируются отростки и у клеток макроглии (астроцитов и олигодендроцитов).
Клетки микроглии образуются из мезенхимы и появляются в ЦНС вместе с прорастанием в нее кровеносных сосудов.
Клетки ганглиозных валиков дифференцируются сначала в биполярные, а затем в псевдоуниполярные чувствительные нервные клетки, центральный отросток которых уходит в ЦНС, а периферический – к рецепторам других тканей и органов, образуя афферентную часть периферической соматической нервной системы.
Эфферентная часть нервной системы состоит из аксонов мотонейронов вентральных отделов нервной трубки.
В первые месяцы постнатального онтогенеза продолжается интенсивный рост аксонов и дендритов и резко возрастает количество синапсов в связи с развитием нейронных сетей.
Эмбриогенез головного мозга начинается с развития в передней (ростральной) части мозговой трубки двух первичных мозговых пузырей, возникающих в результате неравномерного роста стенок нервной трубки (архэнцефалон и дейтерэнцефалон).
Дейтерэнцефалон, как и задняя часть мозговой трубки (впоследствии спинной мозг), располагается над хордой. Архэнцефалон закладывается впереди нее. Затем в начале четвертой недели у зародыша дейтерэнцефалон делится на средний (mesencephalon ) и ромбовидный (rhombencephalon ) пузыри.
А архэнцефалон превращается на этой (трехпузырной) стадии в передний мозговой пузырь (prosencephalon ) (рис.
17, 9 ). В нижней части переднего мозга выпячиваются обонятельные лопасти (из них развиваются обонятельный эпителий носовой полости, обонятельные луковицы и тракты). Из дорсолатеральных стенок переднего мозгового пузыря выступают два глазных пузыря.
В дальнейшем из них развиваются сетчатка глаз, зрительные нервы и тракты.
На шестой неделе эмбрионального развития передний и ромбовидный пузыри делятся каждый на два и наступает пятипузырная стадия (рис. 17).
Передний пузырь – конечный мозг – разделяется продольной щелью на два полушария. Полость также делится, образуя боковые желудочки. Мозговое вещество увеличивается неравномерно, и на поверхности полушарий образуются многочисленные складки – извилины, отделенные друг от друга более или менее глубокими бороздами и щелями (рис.
18). Каждое полушарие разделяется на четыре доли, в соответствие с этим полости боковых желудочков делятся также на 4 части: центральный отдел и три рога желудочка. Из мезенхимы, окружающей мозг зародыша, развиваются оболочки мозга.
Серое вещество располагается и на периферии, образуя кору больших полушарий, и в основании полушарий, образуя подкорковые ядра.

Рис. 18. Этапы развития головного мозга человека
Задняя часть переднего пузыря остается неразделенной и называется теперь промежуточным мозгом (рис.
17, 10 ). Функционально и морфологически он связан с органом зрения. На стадии, когда границы с конечным мозгом слабо выражены, из базальной части боковых стенок образуются парные выросты – глазные пузыри (рис. 17, 8 ), которые соединяются с местом их происхождения при помощи глазных стебельков, впоследствии превращающихся в зрительные нервы. Наибольшей толщины достигают боковые стенки промежуточного мозга, которые преобразуются в зрительные бугры, или таламус.
В соответствии с этим полость III желудочка превращается в узкую сагиттальную щель. В вентральной области (гипоталамус) образуется непарное выпячивание – воронка, из нижнего конца которой происходит задняя мозговая доля гипофиза – нейрогипофиз.
Третий мозговой пузырь превращается в средний мозг (рис.
17, 5), который развивается наиболее просто и отстает в росте. Стенки его утолщаются равномерно, а полость превращается в узкий канал – Сильвиев водопровод, соединяющий III и IV желудочки.
Из дорсальной стенки развивается четверохолмие, а из вентральной – ножки среднего мозга.
Ромбовидный мозг делится на задний и добавочный. Из заднего формируется мозжечок (рис. 17, 12 ) – сначала червь мозжечка, а затем полушария, а также мост (рис. 17, 11 ). Добавочный мозг превращается в продолговатый мозг. Стенки ромбовидного мозга утолщаются – как с боков, так и на дне, только крыша остается в виде тончайшей пластинки.
Полость превращается в IV желудочек, который сообщается с Сильвиевым водопроводом и с центральным каналом спинного мозга.
В результате неравномерного развития мозговых пузырей мозговая трубка начинает изгибаться (на уровне среднего мозга – теменной прогиб, в области заднего мозга – мостовой и в месте перехода добавочного мозга в спинной – затылочный прогиб).
Теменной и затылочный прогибы обращены наружу, а мостовой – внутрь (рис. 17; 18).
Структуры головного мозга, формирующиеся из первичного мозгового пузыря: средний, задний и добавочный мозг – составляют ствол головного мозга (trùncus cere bri ). Он является ростральным продолжением спинного мозга и имеет с ним общие черты строения.
Проходящая по латеральным стенкам спинного мозга и стволового отдела головного мозга парная пограничная борозда (su lcus limitons ) делит мозговую трубку на основную (вентральную) и крыловидную (дорзальную) пластинки. Из основной пластинки формируются моторные структуры (передние рога спинного мозга, двигательные ядра черепно-мозговых нервов).
Над пограничной бороздой из крыловидной пластинки развиваются сенсорные структуры (задние рога спинного мозга, сенсорные ядра ствола мозга), в пределах самой пограничной борозды – центры вегетативной нервной системы.
Производные архэнцефалона (telence phalon и diencéphalon ) создают подкорковые структуры и кору.
Здесь нет основной пластинки (она заканчивается в среднем мозге), следовательно, и нет двигательных и вегетативных ядер.
Весь передний мозг развивается из крыловидной пластинки, поэтому в нем имеются лишь сенсорные структуры (см. рис.18).
Постнатальный онтогенез нервной системы человека начинается с момента рождения ребенка. Головной мозг новорожденного весит 300–400 г. Вскоре после рождения прекращается образование из нейробластов новых нейронов, сами нейроны не делятся. Однако к восьмому месяцу после рождения вес мозга удваивается, а к 4–5 годам утраивается.
Масса мозга растет в основном за счет увеличения количества отростков и их миелинизации. Максимального веса мозг мужчин достигает к 20–29 годам, а женщин к 15–19. После 50 лет мозг уплощается, вес его падает и в старости может уменьшиться на 100 г.
Пермский гуманитарно-технологический институт
Гуманитарный факультет
КОНТРОЛЬНАЯ РАБОТА
по дисциплине «АНАТОМИЯ ЦНС»
на тему
«Основные этапы эволюционного развития ЦНС»
Пермь, 2007
Этапы развития центральной нервной системы
Появление многоклеточных организмов явилось первичным стимулом для дифференциации систем связи, которые обеспечивают целостность реакций организма, взаимодействие между его тканями и органами.
Это взаимодействие может осуществляться как гуморальным путем посредством поступления гормонов и продуктов метаболизма в кровь, лимфу и тканевую жидкость, так и за счет функции нервной системы, которая обеспечивает быструю передачу возбуждения, адресованного к вполне определенным мишеням.
Нервная система беспозвоночных животных
Нервная система как специализированная система интеграции на пути структурного и функционального развития проходит через несколько этапов, которые у первично- и вторичноротых животных могут характеризоваться чертами параллелизма и филогенетической пластичностью выбора.
Среди беспозвоночных наиболее примитивный тип нервной системы в виде диффузной нервной сети встречается у типа кишечнополостных.
Их нервная сеть представляет собой скопление мультиполярных и биполярных нейронов, отростки которых могут перекрещиваться, прилегать друг к другу и лишены функциональной дифференциации на аксоны и дендриты. Диффузная нервная сеть не разделена на центральный и периферический отделы и может быть локализована в эктодерме и энтодерме.
Эпидермальные нервные сплетения, напоминающие нервные сети кишечнополостных, могут быть обнаружены и у более высоко организованных беспозвоночных (плоские и кольчатые черви), однако здесь они занимают соподчиненное положение по отношению к центральной нервной системе (ЦНС), которая выделяется как самостоятельный отдел.
В качестве примера такой централизации и концентрации нервных элементов можно привести ортогональную нервную систему плоских червей.
Ортогон высших турбеллярий представляет собой упорядоченную структуру, которая состоит из ассоциативных и двигательных клеток, формирующих вместе несколько пар продольных тяжей, или стволов, соединенных большим числом поперечных и кольцевых комиссуральных стволов.
Концентрация нервных элементов сопровождается их погружением в глубь тела.
Плоские черви являются билатерально симметричными животными с четко выраженной продольной осью тела. Движение у свободноживущих форм осуществляется преимущественно в сторону головного конца, где концентрируются рецепторы, сигнализирующие о приближении источника раздражения.
К числу таких рецепторов турбеллярий относятся пигментные глазки, обонятельные ямки, статоцист, чувствительные клетки покровов, наличие которых способствует концентрации нервной ткани на переднем конце тела. Этот процесс приводит к формированию головного ганглия, который, по меткому выражению Ч.
Шеррингтона, можно рассматривать как ганглиозную надстройку над системами рецепции на расстоянии.
Ганглионизация нервных элементов получает дальнейшее развитие у высших беспозвоночных, кольчатых червей, моллюсков и членистоногих.
У большинства кольчатых червей брюшные стволы ганглионизированы таким образом, что в каждом сегменте тела формируется по одной паре ганглиев, соединенных коннективами с другой парой, расположенной в соседнем сегменте.
Ганглии одного сегмента у примитивных аннелид соединены между собой поперечными комиссурами, и это приводит к образованию лестничной нервной системы,. В более продвинутых отрядах кольчатых червей наблюдается тенденция к сближению брюшных стволов вплоть до полного слияния ганглиев правой и левой сторон и перехода от лестничной к цепочечной нервной системе. Идентичный, цепочечный тип строения нервной системы существует и у членистоногих с различной выраженностью концентрации нервных элементов, которая может осуществляться не только за счет слияния соседних ганглиев одного сегмента, но и при слиянии последовательных ганглиев различных сегментов.
Эволюция нервной системы беспозвоночных идет не только по пути концентрации нервных элементов, но и в направлении усложнения структурных взаимоотношений в пределах ганглиев.
Не случайно в современной литературе отмечается тенденция сравнивать брюшную нервную цепочку со спинным мозгом позвоночных животных. Как и в спинном мозгу, в ганглиях обнаруживается поверхностное расположение проводящих путей, дифференциация нейропиля на моторную, чувствительную и ассоциативные области.
Это сходство, являющееся примером параллелизма в эволюции тканевых структур, не исключает, однако, своеобразия анатомической организации.
Так, например, расположение туловищного мозга кольчатых червей и членистоногих на брюшной стороне тела обусловило локализацию моторного нейропиля на дорсальной стороне ганглия, а не на вентральной, как это имеет место у позвоночных животных.
Процесс ганглионизации у беспозвоночных может привести к формированию нервной системы разбросанно-узлового типа, которая встречается у моллюсков. В пределах этого многочисленного типа имеются филогенетически примитивные формы с нервной системой, сопоставимой с ортогоном плоских червей (боконервные моллюски), и продвинутые классы (головоногие моллюски), у которых слившиеся ганглии формируют дифференцированный на отделы мозг.
Прогрессивное развитие мозга у головоногих моллюсков и насекомых создает предпосылку для возникновения своеобразной иерархии командных систем управления поведением.
Низший уровень интеграции в сегментарных ганглиях насекомых и в подглоточной массе мозга моллюсков служит основой для автономной деятельности и координации элементарных двигательных актов. В то же время мозг представляет собой следующий, более высокий уровень интеграции, где могут осуществляться межанализаторный синтез и оценка биологической значимости информации.
На основе этих процессов формируются нисходящие команды, обеспечивающие вариантность запуска нейронов сегментарных центров. Очевидно, взаимодействие двух уровней интеграции лежит в основе пластичности поведения высших беспозвоночных, включающего врожденные и приобретенные реакции.
В целом, говоря об эволюции нервной системы беспозвоночных, было бы упрощением представлять ее как линейный процесс.
Факты, полученные в нейроонтогенетических исследованиях беспозвоночных, позволяют допустить множественное (полигенетическое) происхождение нервной ткани беспозвоночных. Следовательно, эволюция нервной системы беспозвоночных могла идти широким фронтом от нескольких источников с изначальным многообразием.
На ранних этапах филогенетического развития сформировался второй ствол эволюционного древа, который дал начало иглокожим и хордовым.
Основным критерием для выделения типа хордовых является наличие хорды, глоточных жаберных щелей и дорсального нервного тяжа - нервной трубки, представляющей собой производное наружного зародышевого листка - эктодермы.
Трубчатый тип нервной системы позвоночных по основным принципам организации отличен от ганглионарного или узлового типа нервной системы высших беспозвоночных.
Нервная система позвоночных животных
Нервная система позвоночных закладывается в виде сплошной нервной трубки, которая в процессе онто- и филогенеза дифференцируется на различные отделы и является также источником периферических симпатических и парасимпатических нервных узлов.
У наиболее древних хордовых (бесчерепных) головной мозг отсутствует и нервная трубка представлена в малодифференцированном состоянии.
Согласно представлениям Л.
А. Орбели, С. Херрика, А. И.
Карамяна, этот критический этап развития центральной нервной системы обозначается как спинальный. Нервная трубка современного бесчерепного (ланцетника), как и спинной мозг более высоко организованных позвоночных, имеет метамерное строение и состоит из 62-64 сегментов, в центре которых проходит спинно-мозговой канал. От каждого сегмента отходят брюшные (двигательные) и спинные (чувствительные) корешки, которые не образуют смешанных нервов, а идут в виде отдельных стволов.
В головных и хвостовых отделах нервной трубки локализованы гигантские клетки Родэ, толстые аксоны которых образуют проводниковый аппарат. С клетками Родэ связаны светочувствительные глазки Гесса, возбуждение которых вызывает отрицательный фототаксис.
В головной части нервной трубки ланцетника находятся крупные ганглиозные клетки Овсянникова, имеющие синаптические контакты с биполярными чувствительными клетками обонятельной ямки.
В последнее время в головной части нервной трубки идентифицированы нейросекреторные клетки, напоминающие гипофизарную систему высших позвоночных. Однако анализ восприятия и простых форм обучения ланцетника показывает, что на данном этапе развития ЦНС функционирует по принципу эквипотенциальности, и утверждение о специфике головного отдела нервной трубки не имеет достаточных оснований.
В ходе дальнейшей эволюции наблюдается перемещение некоторых функций и систем интеграции из спинного мозга в головной - процесс энцефализации, который был рассмотрен на примере беспозвоночных животных.
В период филогенетического развития от уровня бесчерепных до уровня круглоротых формируется головной мозг как надстройка над системами дистантной рецепции.
Исследование ЦНС современных круглоротых показывает, что их головной мозг в зачаточном состоянии содержит все основные структурные элементы.
Развитие вестибулолатеральной системы, связанной с полукружными каналами и рецепторами боковой линии, возникновение ядер блуждающего нерва и дыхательного центра создают основу для формирования заднего мозга. Задний мозг миноги включает продолговатый мозг и мозжечок в виде небольших выпячиваний нервной трубки.
Общее развитие нервной системы
Филогенез нервной системы в кратких чертах сводится к следующему. У простейших одноклеточных организмов (амеба) нервной системы еще нет, а связь с окружающей средой осуществляется при помощи жидкостей, находящихся внутри и вне организма - гуморальная (humor - жидкость), донервная форма регуляции.
В дальнейшем, когда возникает нервная система, появляется и другая форма регуляции - нервная.
По мере развития нервной системы нервная регуляция все больше подчиняет себе гуморальную, так что образуется единая нейро-гуморальная регуляция при ведущей роли нервной системы. Последняя в процессе филогенеза проходит ряд основных этапов (рис.

I этап - сетевидная нервная система. На этом этапе (кишечнополостные) нервная система, например гидры, состоит из нервных клеток, многочисленные отростки которых соединяются друг с другом в разных направлениях, образуя сеть, диффузно пронизывающую все тело животного.
При раздражении любой точки тела возбуждение разливается по всей нервной сети, и животное реагирует движением всего тела. Отражением этого этапа у человека является сетевидное строение интрамуральной нервной системы.
II этап - узловая нервная система.
На этом этапе (высшие черви) нервные клетки сближаются в отдельные скопления или группы, причем из скоплений клеточных тел получаются нервные узлы - центры, а из скоплений отростков - нервные стволы - нервы. При этом в каждой клетке число отростков уменьшается, и они получают определенное направление.
Соответственно сегментарному строению тела животного, например у кольчатого червя, в каждом сегменте имеются сегментарные нервные узлы и нервные стволы. Последние соединяют узлы в двух направлениях поперечные стволы связывают узлы данного сегмента, а продольные - узлы разных сегментов.
Благодаря этому нервные импульсы, возникающие в какой-либо точке тела, не разливаются по всему телу, а распространяются по поперечным стволам в пределах данного сегмента.
Продольные стволы связывают нервные сегменты в одно целое. На головном конце животного, который при движении вперед соприкасается с различными предметами окружающего мира, развиваются органы чувств, в связи с чем головные узлы развиваются сильнее остальных, являясь прообразом будущего головного мозга.
Отражением этого этапа является сохранение у человека примитивных черт (разбросанность на периферии узлов и микроганглиев) в строении вегетативной нервной системы.
III этап - трубчатая нервная система. На первоначальной ступени развития животных особенно большую роль играл аппарат движения, от совершенства которого зависит основное условие существования животного - питание (передвижение в поисках пищи, захватывание и поглощение ее).
У низших многоклеточных развился перистальтический способ передвижения, что связано с гладкой мускулатурой и ее местным нервным аппаратом.
На более высокой ступени перистальтический способ сменяется скелетной моторикой, т. е. передвижением с помощью системы жестких рычагов - поверх мышц (членистоногие) и внутри мышц (позвоночные).
Следствием этого явилось образование поперечнополосатой мускулатуры и центральной нервной системы, координирующей перемещение отдельных рычагов моторного скелета.
Такая центральная нервная система у хордовых (ланцетник) возникла в виде метамерно построенной нервной трубки с отходящими от нее сегментарными нервами ко всем сегментам тела, включая и аппарат движения - туловищный мозг.
У позвоночных и человека туловищный мозг становится спинным. Таким образом, появление туловищного мозга связано с усовершенствованием в первую очередь моторного вооружения животного.
Наряду с этим уже у ланцетника имеются и рецепторы (обонятельный, световой). Дальнейшее развитие нервной системы и возникновение головного мозга обусловлены преимущественно усовершенствованием рецепторного вооружения.
Так как большинство органов чувств возникает на том конце тела животного, который обращен в сторону движения, т. е. вперед, то для восприятия поступающих через них внешних раздражений развивается передний конец туловищного мозга и образуется головной мозг, что совпадает с обособлением переднего конца тела в виде головы - цефализация (cephal - голова).
К. Сепп в руководстве по нервным болезням дает упрощенную, но удобную для изучения схему филогенеза головного мозга, которую мы и приводим. Согласно этой схеме, на первом этапе развития головной мозг состоит из трех отделов: заднего, среднего и переднего, причем из этих отделов в первую очередь (у низших рыб) особенно развивается задний, или ромбовидный, мозг (rhombencephalon).
Развитие заднего мозга происходит под влиянием рецепторов акустики и статики (рецепторы VIII пары головных нервов), имеющих ведущее значение для ориентации в водной среде.
В дальнейшей эволюции задний мозг дифференцируется на продолговатый мозг, являющийся переходным отделом от спинного мозга к головному и потому называемый myelencephalon (myelos - спинной мозг, encephalon - головной), и собственно задний мозг - metencephalon, из которого развиваются мозжечок и мост.
В процессе приспособления организма к окружающей среде путем изменения обмена веществ в заднем мозгу как наиболее развитом на этом этапе отделе центральной нервной системы возникают центры управления жизненно важными процессами растительной жизни, связанными, в частности, с жаберным аппаратом (дыхание, кровообращение, пищеварение и др.).
Поэтому в продолговатом мозгу возникают ядра жаберных нервов (группа X пары - вагуса). Эти жизненно важные центры дыхания и кровообращения остаются в продолговатом мозгу человека, чем объясняется смерть, наступающая при повреждении продолговатого мозга. На II этапе (еще у рыб) под влиянием зрительного рецептора особенно развивается средний мозг, mesencephalon. На III этапе, в связи с окончательным переходом животных из водной среды в воздушную, усиленно развивается обонятельный рецептор, воспринимающий содержащиеся в воздухе химические вещества, сигнализирующие своим запахом о добыче, опасности и других жизненно важных явлениях окружающей природы.
Под влиянием обонятельного рецептора развивается передний мозг — prosencephalon, вначале имеющий характер чисто обонятельного мозга.
В дальнейшем передний мозг разрастается и дифференцируется на промежуточный - diencephalon и конечный - telencephalon.
В конечном мозгу как в высшем отделе центральной нервной системы появляются центры для всех видов чувствительности. Однако нижележащие центры не исчезают, а сохраняются, подчиняясь центрам вышележащего этажа. Следовательно, с каждым новым этапом развития головного мозга возникают новые центры, подчиняющие себе старые.
Происходит как бы передвижение функциональных центров к головному концу и одновременное подчинение филогенетически старых зачатков новым. В результате центры слуха, впервые возникшие в заднем мозгу, имеются также в среднем и переднем, центры зрения, возникшие в среднем, имеются и в переднем, а центры обоняния - только в переднем мозгу.
Под влиянием обонятельного рецептора развивается небольшая часть переднего мозга, называемая поэтому обонятельным мозгом (rhinencephalon), который покрыт корой серого вещества - старой корой (paleocortex).
Совершенствование рецепторов приводит к прогрессивном;, развитию переднего мозга, который постепенно становится органом, управляющим всем поведением животного.
Различают две формы поведения животного: инстинктивное, основанное на видовых реакциях (безусловные рефлексы), и индивидуальное, основанное на опыте индивида (условные рефлексы).
Соответственно этим двум формам поведения в конечном мозгу развиваются две группы центров серого вещества: подкорковые узы, имеющие строение ядер (ядерные центры), и кора серого вещества, имеющая строение сплошного экрана (экранные центры). При этом вначале развивается «подкорка», а затем кора. Кора возникает при переходе животного от водного к наземному образу жизни и обнаруживается отчетливо у амфибий и рептилий.
Дальнейшая эволюция нервной системы характеризуется тем, что кора головного мозга все более и более подчиняет себе функции всех нижележащих центров, происходит постепенная кортиколизация функций.
Пневматоз желудка
Необходимой формацией для осуществления высшей нервной деятельности является новая кора, расположенная на поверхности полушарий и приобретающая в процессе филогенеза шестислойное строение.
Благодаря усиленному развитию новой коры конечный мозг у высших позвоночных превосходит все остальные отделы головного мозга, покрывая их, как плащом (pallium). Развивающийся новый мозг (neencephalon) оттесняет в глубину старый мозг (обонятельный), который как бы свертывается в виде аммонова рога (cornu Ammoni или pes hyppocampi), остающегося по-прежнему обонятельным центром. В результате плащ, т. е. новый мозг (neencephalon), резко преобладает над остальными отделами мозга - старым мозгом (paleencephalon).
Итак, развитие головного мозга совершается под влиянием развития рецепторов, чем и объясняется, что самый высший отдел головного мозга - кора серого вещества - представляет, как учит И.
П. Павлов, совокупность корковых концов анализаторов, т. е. сплошную воспринимающую (рецепторную) поверхность.
Дальнейшее развитие мозга у человека подчиняется иным закономерностям, связанным с его социальной природой. Кроме естественных органов тела, имеющихся и у животных, человек стал пользоваться орудиями труда.
Орудия труда, ставшие искусственными органами, дополнили естественные органы тела и составили техническое вооружение человека.
С помощью этого вооружения человек приобрел возможность не только приспосабливаться самому к природе, как это делают животные, но и приспосабливать природу к своим нуждам.
Труд, как уже говорилось выше, явился решающим фактором становления человека, а в процессе общественного труда возникло необходимое для общения людей средство - речь. «Сначала труд, а затем и вместе с ним членораздельная речь явились двумя самыми главными стимулами, под влиянием которых мозг обезьяны постепенно превратился в человеческий мозг, который, при всем своем сходстве с обезьяньим, далеко превосходит его по величине и совершенству».
Это совершенство обусловлено максимальным развитием конечного мозга, особенно его коры - новой коры (neocortex).
Кроме анализаторов, воспринимающих различные раздражения внешнего мира и составляющих материальный субстрат конкретно-наглядного мышления, свойственного животным (первая сигнальная система действительности,), у человека возникла способность асбстрактного, отвлеченного мышления с помощью слова, сначала слышимого (устная речь) и позднее видимого (письменная речь).
Это составило вторую сигнальную систему, по И. П. Павлову, которая в развивающемся животном мире явилась «чрезвычайной прибавкой к механизмам нервной деятельности». Материальным субстратом второй сигнальной системы стали поверхностные слои новой коры. Поэтому кора конечного мозга достигает своего наивысшего развития у человека.
Таким образом, вся эволюция нервной системы сводится к прогрессивному развитию конечного мозга, который у высших позвоночных и особенно у человека в связи с усложнением нервных функций достигает огромных размеров.
Изложенные закономерности филогенеза обусловливают эмбриогенез нервной системы человека. Нервная система происходит из наружного зародышевого листка, или эктодермы. Эта последняя образует продольное утолщение, называемое медуллярной пластинкой (рис.

Медуллярная пластинка скоро углубляется в медулляярную бороздку, края которой (медуллярные валики) постепенно становятся выше и затем срастаются друг с другом, превращая бороздку в трубку (мозговая трубка).
Мозговая трубка представляет собой зачаток центральной части нервной системы. Задний конец трубки образует зачаток спинного мозга, передний же ее расширенный конец путем перетяжек расчленяется на три первичных мозговых пузырька, из которых происходит головной мозг во всей его сложности.
Мозговая пластинка первоначально состоит только из одного слоя эпителиальных клеток.
Во время замыкания ее в мозговую трубку количество клеток в стенках последней увеличивается, так что возникает три слоя: внутренний (обращенный в полость трубки), из которого происходит эпителиальная выстилка мозговых полостей (эпендима центрального канала спинного мозга и желудочков головного); средний, из которого развивается серое вещество мозга (нервные клетки - нейробласты), наконец, наружный, почти не содержащий клеточных ядер, развивающийся в белое вещество (отростки нервных клеток - нейриты).
Пучки нейритов нейробластов распространяются или в толще мозговой трубки, образуя белое вещество мозга, или же выходят в мезодерму и затем соединяются с молодыми мышечными клетками (миобластами). Таким путем возникают двигательные нервы.
Чувствительные нервы возникают из зачатков спинномозговых узлов, которые заметны уже по краям медуллярной бороздки у места перехода ее в кожную эктодерму. Когда бороздка смыкается в мозговую трубку, зачатки смещаются на ее дорсальную сторону, располагаясь по средней линии.
Затем клетки этих зачатков перемещаются вентрально и располагаются вновь по бокам мозговой трубки в виде так называемых ганглиозных валиков. Оба ганглиозных валика перешнуровываются четкообразно по сегментам дорсальной стороны зародыша, вследствие чего получается на каждой стороне ряд спинномозговых узлов, ganglia spinalia s.
intervertebral. В головной части мозговой трубки они доходят только до области заднего мозгового пузырька, где образуют зачатки узлов чувствительных головных нервов. В ганглиозных зачатках развиваются нейробласты, принимающие вид биполярных нервных клеток, один из отростков которых врастает в мозговую трубку, другой идет на периферию, образуя чувствительный нерв. Благодаря сращению на некотором протяжении от начала обоих отростков получаются из биполярных так называемые ложные униполярные клетки с одним отростком, делящимся в форме буквы «Т», являющиеся характерными для межпозвонковых узлов взрослого.
Центральные отростки клеток, проникающие в спинной мозг, составляют задние корешки спинномозговых нервов, а периферические отростки, разрастаясь вентрально, образуют (вместе с вышедшими из спинного мозга эфферентными волокнами, составляющими передний корешок) смешанный спинномозговой нерв.
Онтогенез (оntogenesis; греч. оп, ontos - сущее + genesis - зарождение, происхождение) - процесс индивидуального развития организма от момента его зарождения (зачатия) до смерти. Выделяют: эмбриональный (зародышевый, или пренатальный) — время от оплодотворения до рождения и постэмбриональный (послезародышевый, или постнатальный) — от рождения до смерти, периоды развития.
Нервная система человека развивается из эктодермы — наружного зародышевого листка.
В конце второй недели эмбрионального развития в дорсальных отделах туловища обособляется участок эпителия – нервная (медуллярная) пластинка , клетки которой интенсивно размножаются и дифференцируются. Ускоренный рост боковых отделов нервной пластинки приводит к тому, что ее края сначала приподнимаются, затем сближаются и, наконец, в конце третьей недели срастаются, формируя первичную мозговую трубку .
После чего мозговая трубка постепенно погружается в мезодерму.
Рис.1. Формирование нервной трубки.

Нервная трубка представляет собой эмбриональный зачаток всей нервной системы человека.
Из нее в дальнейшем формируется головной и спинной мозг, а также периферические отделы нервной системы. При смыкании нервного желобка по бокам в области его приподнятых краев (нервных валиков) с каждой стороны выделяется группа клеток, которая по мере обособления нервной трубки от кожной эктодермы образует между нервными валиками и эктодермой сплошной слой - ганглиозную пластинку.
Последняя служит исходным материалом для клеток чувствительных нервных узлов (спинно- и черепномозговых ганглиев) и узлов вегетативной нервной системы, иннервирующей внутренние органы.
Нервная трубка на ранней стадии своего развития состоит из одного слоя клеток цилиндрической формы, которые в дальнейшем интенсивно размножаются митозом и количество их увеличивается; в результате стенка нервной трубки утолщается.
В этой стадии развития в ней можно выделить три слоя: внутренний (в дальнейшем из него сформируется эпендимальная выстилка), среднего слоя (серое вещество мозга, клеточные элементы этого слоя дифференцируются в двух направлениях: часть их превращается в нейроны, другая часть - в глиальные клетки) и наружного слоя (белое вещество мозга).
Рис.2 .
Этапы развитияголовного мозга человека.

Нервная трубка развивается неравномерно. Вследствие интенсивного развития ее передней части начинает формироваться головной мозг, образуются мозговые пузыри: вначале появляются два пузыря, затем задний пузырь делится еще на два. В результате у четырехнедельных эмбрионов головной мозг состоит из трех мозговых пузырей (передний, средний и ромбовидный мозг).
На пятой неделе передний мозговой пузырь подразделяется на конечный мозг и промежуточный, а ромбовидный – на задний и продолговатый (стадия пяти мозговых пузырей ). Одновременно нервная трубка образует несколько изгибов в сагитальной плоскости.
Из недифференцированной задней части медуллярной трубки развивается спинной мозг со спинно-мозговым каналом. Из полостей эмбрионального головного мозга происходит формирование желудочков мозга .
Полость ромбовидного мозга преобразуется в IY желудочек, полость среднего мозга формирует водопровод мозга, полость промежуточного мозга образует III желудочек мозга, а полость переднего мозга — имеющие сложную конфигурацию боковые желудочки мозга.
После формирования пяти мозговых пузырей в структурах нервной системы происходят сложные процессы внутренней дифференцировки и роста различных отделов мозга.
На 5-10 неделе наблюдается рост и дифференцировка конечного мозга: образуются корковые и подкорковые центры, происходит расслоение коры. Образуются мозговые оболочки. Спинной мозг приобретает дефинитивное состояние. На 10-20 неделе завершаются процессы миграции, формируются все основные отделы головного мозга, на первый план выходят процессы дифференцировки.
Наиболее активно развивается конечный мозг. Полушария головного мозга становятся самой крупной частью нервной системы. На 4-м месяце развития плода человека появляется поперечная щель большого мозга, на 6-м - центральная борозда и другие главные борозды, в последующие месяцы - второстепенные и после рождения - самые мелкие борозды.
В процессе развития нервной системы важную роль играет миелинизация нервных волокон, в результате которой нервные волокна покрываются защитным слоем миелина и значительно вырастает скорость проведения нервных импульсов.
К концу 4-го месяца внутриутробного развития миелин выявляется в нервных волокнах, составляющих восходящие, или афферентные (чувствительные), системы боковых канатиков спинного мозга, тогда как в волокнах нисходящих, или эфферентных (двигательных), систем миелин обнаруживается на 6-м месяце.
Приблизительно в это же время наступает миелинизация нервных волокон задних канатиков. Миелинизация нервных волокон корково-спинномозговых путей начинается на последнем месяце внутриутробной жизни и продолжается в течение года после рождения.
Это свидетельствуются о том, что процесс миелинизации нервных волокон распространяется вначале на филогенетически более древние, а затем - на более молодые структуры. От последовательности миелинизации определенных нервных структур зависит очередность формирования их функций.
Формирование функции и также зависит и от дифференциации клеточных элементов и их постепенного созревания, которое длится в течение первого десятилетия.
К моменту рождения ребенка нервные клетки достигают зрелости и уже неспособны к делению. В связи с этим в дальнейшем их число не увеличивается.
В постнатальном периоде постепенно происходит окончательное созревание всей нервной системы, в частности ее самого сложного отдела - коры большого мозга, играющей особую роль в мозговых механизмах условно-рефлекторной деятельности, формирующейся с первых дней жизни.
Еще один важный этап в онтогенезе это период полового созревания, когда проходит и половая дифференцировка мозга.
В течение всей жизни человека мозг активно изменяется, приспосабливаясь к условиям внешней и внутренней среды, часть этих изменений носит генетически запрограммированный характер, часть является относительно свободной реакцией на условия существования. Онтогенез нервной системы заканчивается только со смертью человека.